
Our latest paper on the acute effects of nicotinamide mononucleotide (NMN) is out in J. Mol. Cell. Cardiol. For those without access to the journal, a pre-print of the paper was placed in BioRxiv and is still there.
The paper stems from some work we started a few years ago, trying to find out if the lysine deacetylase SIRT1 can regulate intracellular pH. In a previous paper, we had shown that the SIRT1 inhibitor splitomicin can cause cytosolic alkalinization in cardiomyocytes (Figure 4 of that paper). So, the next logical step was to see if stimulating SIRT1 can do the opposite. In addition, we’d published several papers showing that SIRT1 activation is cardioprotective against ischemia-reperfusion injury, so we had a general interest in seeing if SIRT1 stimulators might be protecting the heart by altering pH.
Behold, NAD+ supplements!
There’s a giant poo-storm in the literature about how best to stimulate SIRT1, with resveratrol being everyone’s least favorite molecule, and other alleged compounds (e.g., the Sirtris molecules) being of questionable efficacy. But, many people have been paying attention to the growing field of NAD+ supplementation. Since the sirtuin enzymes use NAD+ as a substrate, there’s a lot of interest in orally bioavailable precursors of NAD+, including nicotinamide mononucleotide (NMN) and nicotinamide riboside (NR).
Naturally, with any small molecule that has potential ability to prevent or reverse diseases of aging, there are significant commercial interests at play. Namely, Elysium (Leonard Guarente and David Sinclair) is shilling NR pills under the name “Basis”, at $40 for a month’s supply. Chromadex is selling a similar product under the name “Tru-Niagen” at $50. Other companies are selling NMN with all sorts of health claims, and if you’re cheap like me you can even buy NMN on Amazon. Naturally, the mainstream press is beginning to pay attention to all this fountain of youth chatter.
The data…
So, what happened when we put NMN into hearts and heart cells? It acidified the ever-loving crap out of them! Here are the key data, using a fluorescent pH indicator in adult mouse primary cardimyocytes…

So all good right? Hypothesis proven – stimulating SIRT1 makes cells acid, just as inhibiting it makes them alkaline. Not to just figure out how SIRT1 is regulating pH and we’re all set. Not so fast! It turned out that blocking SIRT1 didn’t do anything to NMN’s ability to acidify, so it must have been working via another mechanism.
Long story short – the mechanism is simple bulk stimulation of glycolysis. As you will recall from grade school biochemistry, a key step in glycolysis is GAPDH, which uses NAD+. The reason we have lactate dehydrogenase (LDH) is to convert the NADH generated by GAPDH back into NAD+, so the pathway can continue working. Ergo, without NAD+ glycolysis cannot work, and so boosting NAD+ would be predicted to make it work faster…

(If you don’t know this stuff by heart, do yourself a favor and grab a copy of the excellent “Metabolism at a Glance” by J.G. Salway. It’s a required desktop accessory in any serious metabolism/mitochondria lab). Anyway, using various methods including 13C-glucose flux tracing, we showed that NMN boosts glycolysis.
We also looked at lysine acetylation, as a surrogate marker for sirtuin activity (since the SIRT enzymes are lysine deacetylases). In a quite surprising result, we also found that NMN caused robust deacetylation inside mitochondria, without any impact in the cytosol. 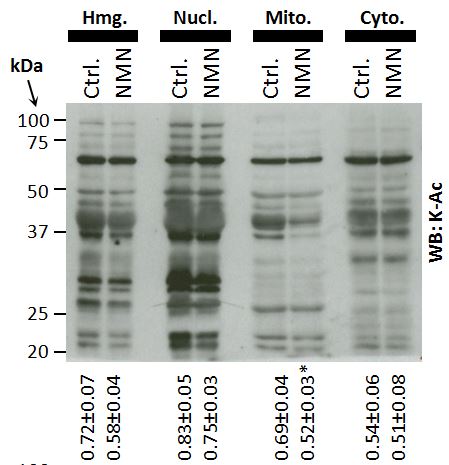
A probable explanation for this is that SIRT1 (which is mostly cytosolic in myocytes) doesn’t really get a look-in on the bolus of NAD+ made from NMN. There’s an crap-ton of GAPDH around (after all, what do we all use as a housekeeping marker protein on our western blots?), so it just chews through all the NAD+, depriving SIRT1 of that sweet nucleotide action. We assume that in the mito’, the NAD+ pool is protected, and so deacetylation can still occur via mito’ sirtuins such as SIRT3. Some excellent recent work by Joe Baur on NAD compartmentalization and synthesis is probably relevant to this point.
We also know from some work done by Jim Downey and others that acidic pH in the heart can be protective in the setting of reperfusion injury (acid pH keeps the mito’ permeability transition pore closed). So, we were able to show that delivering a burst of acidosis by giving NMN at the moment of reperfusion is also cardioprotective. We also showed that blocking glycolysis (by removing glucose from the media or by using galactose to prevent glycolysis from making any net ATP) can block the cardioprotective effects of NMN. Overall, although it had already been shown that NMN is cardioprotective, the mechanism was thought to involve SIRT1, but our work now shows that glycolysis and acid pH can also play a role.
So, what does this all mean for the burgeoning NAD+ supplementation industry?
First off, I should stress that the effects of NMN we saw here were all acute, taking place within 10-30 minutes of addition. Nevertheless, I think there are some implications for the human usage of NMN or NR supplements….
(i) The Warburg effect is a key feature of cancer. To put it in simple terms, cancer cells are addicted to glycolysis. There’s also some literature suggesting that stem cells (which share some metabolic features with cancer cells) can benefit from NAD+ supplementation. So, what do you think would happen if we acutely boosted glycolysis in cancer cells? Is it wise to boost NAD+ levels in cancer patients?
(ii) Although the overall tone of this post might seem negative toward the entire field of NAD+ supplementation, it’s hard to argue with the rather impressive effects that have so far been reported in animal models and even in humans. As such, it’s worth asking whether acute, transient metabolic acidosis might actually be part of the beneficial mechanism of these molecules? On that topic, there’s another common intervention that causes acute metabolic acidosis, and is rather famous for having beneficial effects in various diseases… exercise!
Evolving open publication strategies
As suggested in the post title, it’s worth highlighting that this is now the 6th paper for which we’ve posted a pre-print on BioRxiv. A couple of years ago when we dipped a toe in the pre-print water for the first time (with this paper), it was with some trepidation! The surprise for me is how this has now become the standard publication workflow for my lab – we just post pre-prints without really thinking about it. Similarly, almost 3 years ago for the first time we published the complete raw data set for a paper on FigShare. Now it’s just something we do as part of the normal paper preparation process. Sure, it’s a lot of work to curate everything into an accessible format, but it also really makes you look at your data carefully, so is helpful in preventing mistakes. As a bonus – mentioning both of these strategies (pre-prints and open data) in the Rigor & Reproducibility section of the NIH grant proposal forms has so far worked very much in our favor! It’s a tangible way to show reviewers that you’re walking-the-walk, not just talking-the-talk.